
N'oubliez pas que vous pouvez vous abonner aux nouvelles de cette page grâce à ce flux rss : 
Les anciennes actualités sont déplacées au fur et à mesure dans les archives.

Bientôt mis à jour
Le régime des ammonites (28/2/2011)
Kruta I., Landman N., Rouget I., Cecca F. and Tafforeau P., 2011. The role of ammonites in the Mesozoic marine food web revealed by jaw preservation. Science 331(6013): 70-72
Apparues au Dévonien, il y a 407 Ma, les ammonites forment un groupe de mollusques céphalopodes très prisé par les amateurs de fossiles. Durant 340 Ma, elles se sont diversifiées, souvent de manière très rapide, constituant un sujet d'étude également privilégié par les paléontologues, notamment pour la biostratigraphie. Leur groupe s'est éteint il y a 65 Ma, lors de la crise Crétacé-Tertiaire. On les compare souvent au nautile, bien qu'elles soient plus proches des calmars et des pieuvres. Grâce au synchrotron, une équipe a pu obtenir des images très nettes de l'appareil buccal de Baculites (un genre à la coquille droite), ammonites du Crétacé supérieur des Etats-Unis. Celui-ci est composé de 2 mâchoires et d'une radula chitineuses. Chez l'un des spécimens, les restes de son dernier repas ont également pu être identifiés : des morceaux d'isopode (un groupe de Crustacés, comprenant notamment les cloportes) et la coquille larvaire d'un gastéropode benthique (la larve était pélagique par contre). Ces proies nageaient dans la colonne d'eau, prenant part au plancton. Les chercheurs se sont intéressés à la morphologie des pièces buccales, les comparant avec celles de mollusques actuels, afin d'apporter des précisions quant au régime alimentaire, encore peu connu, des ammonites. En effet, on a longtemps inféré un mode de vie proche de celui des nautiles, qui sont des carnassiers et des charognards. Les mâchoires de Baculites étaient cependant trop fragiles et trop peu abîmées pour avoir mordu et dépecé de grosses proies. Au contraire, elles sont semblables à celles des mollusques actuels se nourrissant de proies petites ou molles. La morphologie de l'appareil buccal et les fragments de proie impliquent que Baculites se nourrissait de plancton; ces ammonites n'auraient d'ailleurs pas été capables de dépecer et d'ingérer des proies plus importantes.
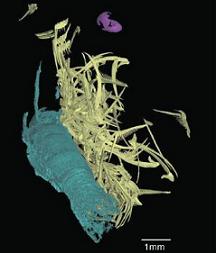
Reconstruction d'une partie de l'appareil buccal de Baculites (spécimen AMNH 66253); la radula est en jaune, le fragment d'isopode en bleu, et la coquille larvaire de gastéropode en mauve (Figure issue de Kruta et al., 2011)
Leur diversification peut ainsi être corrélée à celle du plancton au cours du Jurassique inférieur. De même, lorsque ce dernier fut sévèrement touché lors de la crise Crétacé-Tertiaire, les ammonites se sont éteintes. Le fait que toutes les ammonites aient disparu il y a 65 Ma, y compris les carnivores, alors que les nautiles qui avaient un mode de vie semblable ont survécu, n'est pas encore compris. Il est possible que les larves d'ammonites étant plus petites que celles de nautiles, elles dépendaient plus du plancton lors des premiers stades de vie. Le déclin abrupt du plancton les aurait ainsi plus durement touchées, éradiquant le groupe.