
L'odorat chez les oiseaux
Contrairement à ce que l'on pourrait croire, les oiseaux ne sont pas dépourvus d'odorat. Il y a bien sûr les vautours (et particulièrement l'urubu à tête rouge), dont les capacités olfactives sont connues depuis longtemps. Egalement le kiwi, oiseau aptère de Nouvelle-Zélande, dont l'une des particularités est d'avoir les narines situées à l'extrémité du bec. Leur odorat fin leur permet de détecter les vers et insectes dont ils se nourrissent en fouissant le sol de leur long bec. On pensait que l'odorat des autres oiseaux était rudimentaire, voire inexistant.

Kiwi roa (© Gerald Cubitt / www.nhpa.co.uk)
Par quelques observations fortuites, ainsi que le fait que certains oiseaux exhalent une odeur forte (comme les stariques (cousins des pingouins et guillemots) ou le kakapo (perroquet de Nouvelle-Zélande)), on présumait donc que certains oiseaux étaient doués d'odorat. Il était temps de mettre en place quelques expériences rigoureuses pour étayer ces observations.
Plusieurs expériences ont été réalisées, la plus classique étant celle où un oiseau doit choisir entre deux chemins (aux odeurs différentes). Dans l'une d'elles, des roselins familiers allaient chercher leur nourriture dans un flux d'air qui passait auparavant par des excréments de chat, de lapin ou de l'eau. On a observé que les oiseaux passaient moins de temps auprès de la nourriture lorsque l'air était chargé d'une odeur d'excrément, l'effet étant plus marqué pour les excréments de chat. Cette expérience tend à montrer que ces oiseaux peuvent détecter la présence de prédateurs à l'odeur de leurs excréments.
D'autres chercheurs se sont penchés sur le génome des oiseaux, et ont cherché des gènes spécifiques à l'odorat : ceux des récepteurs olfactifs. Ces gènes sont exprimés dans les neurones sensoriels de l'épithélium olfactif. Leur nombre est un indicateur du nombre d'odeurs différentes qu'une espèce peut discriminer. De grandes différences dans le nombre de ces gènes ont été observés selon les espèces d'oiseau (l'échantillon recouvrait plusieurs familles différentes). Sans surprise, c'est le kiwi qui en exhibe le plus grand nombre; il en possède environ 6 fois plus que le canari. La plupart de ces gènes sont fonctionnels chez les oiseaux, indiquant encore une fois l'importance insoupçonnée de l'odorat chez eux (chez l'homme, qui a un sens de l'odorat peu développé, seuls 40% des gènes des récepteurs olfactifs sont fonctionnels; les autres ont accumulé des mutations et ne sont plus utilisables). L'analyse des génomes aviens a même permis de mettre en évidence une nouvelle classe de gènes de récepteurs olfactifs; celle-ci semble partagée par tous les oiseaux, tout en étant absente chez les autres vertébrés.
Un cas en particulier : les oiseaux de mer
Penchons-nous maintenant sur un groupe qui a fait l'objet d'études plus poussées, les Procellariiformes (voir références ci-dessous). Les Procellariiformes regroupent notamment les pétrels, albatros, fulmars, prions et puffins. Ces oiseaux marins passent la majeure partie de leur vie en mer, ne revenant à terre que quelques mois par an pour s'accoupler et élever leur jeune. Ils parcourent les mers à la recherche de leurs proies : krill, poissons et calmars. Ils forment des couples stables et fidèles à leur nid. Ils sont connus pour leur odeur forte et musquée, qui imprègne leur plumage et leur nid. Ils ont également des bulbes olfactifs très larges.
Les proies des Procellariiformes sont souvent regroupées en bancs mais leur distribution dans l'espace est assez inégale. Ces oiseaux possèdent deux techniques de chasse différentes pour pallier à cet inconvénient. Soit d'une manière dite opportuniste (comme l'albatros hurleur) : en parcourant des milliers de km sous forme de boucles, l'animal rencontre au hasard des zones abondantes en proies. Soit comme un "navetteur" (tel l'albatros à sourcils noirs), où il s'agit pour l'oiseau de rejoindre une zone où les proies sont généralement plus concentrées; à partir de là il s'engage dans une recherche restreinte à cette zone (ARS, pour area-restricted search) pour localiser ses proies.
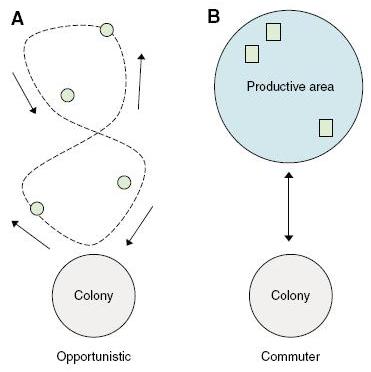
A) Stratégie opportuniste, B) stratégie commutative (Photo issue de Nevitt, 2008)
Les reliefs sous-marins rassemblent souvent du phytoplancton, qui nourrit toute une série d'animaux qui sont eux-même les proies des oiseaux. Nevitt propose que ces zones ont une odeur particulière, et que les oiseaux s'en servent pour les reconnaître et trouver leurs proies, avec éventuellement d'autres indices tels que la vue et l'observation des activités menées par d'autres oiseaux dans la zone. La molécule-clé serait le sulfure de diméthyle (DMS) ou méthylthiométhane, un composé issu de la décomposition métabolique du diméthylsulfoniopropionate chez le phytoplancton marin et autres algues lorsqu'elles sont écrasées, décomposition qui se fait via les processus de la chaîne alimentaire microbienne. Dans l'Océan Antarctique, le DMS est souvent associé à des zones où le plancton est abondant, et où les prédateurs marins ont tendance à s'aggréger.
Les oiseaux ne sont donc pas attirés par l'odeur directe des proies, mais par des odeurs résultant des activités de nourrissage de celles-ci, telles celle du DMS. Lors d'expériences, on a montré que plusieurs espèces d'oiseaux marins répondent à ces odeurs en remontant jusqu'à leur source par un comportement de recherche en zig-zag, contre le vent. Le DMS est détecté par les oiseaux jusqu'à des concentrations de l'ordre de 10-12 mol/l. Ces mêmes espèces ignoraient par contre l'odeur du krill, qui est pourtant leur principale nourriture. Les oiseaux suivent aussi les odeurs de poisson, probablement car celles-ci indiquent les navires de pêche auprès desquels ils aiment à se nourrir.
Les poussins des oiseaux de mer montrent très tôt une attirance pour les odeurs liées aux proies, avant même d'avoir quitté le nid (qui chez certaines espèces est même un terrier). Cette attirance est probablement apprise par leurs parents, qui sentent souvent le DMS ou le phytoplancton lorsqu'ils reviennent nourrir le poussin. Celui-ci associe alors ces odeurs à celle de la nourriture, ce qui au moment de l'envol pourrait l'aider à trouver des sites de nourrissage potentiels. Les poussins sont abrités dans des terriers pendant plusieurs semaines avant leur envol et vivent dans un environnement sombre où les odeurs sont parmi leurs premières expériences sensorielles. Les espèces qui élèvent leurs jeunes dans un terrier sont aussi celles qui montrent un comportement de recherche vis-à-vis du DMS. Les sites profitables pour se nourrir en mer sont dynamiques, c'est-à-dire qu'ils sont fort susceptibles de changer dans l'espace et dans le temps. Les prédateurs ne peuvent donc pas rester cantonnés à une zone de pêche mais doivent sans cesse chercher leurs proies à des endroits différents.

Albatros adulte et son poussin (© www.newworldencyclopedia.org)
Des expériences menées sur des poussins et des adultes de pétrels et prions ont montré qu'ils se servent essentiellement de leur odorat pour retrouver le nid lorsqu'ils en sont éloignés. Ils sont également capables de faire la différence entre leur propre odeur et celle de leur partenaire ou d'un autre membre de leur colonie.
L'odorat chez les oiseaux ne servirait donc pas uniquement à la recherche de nourriture mais pourrait aussi être employé dans le choix d'un partenaire sexuel par exemple. Etant donné que ces oiseaux forment des couples stables et fidèles, avec un cycle de reproduction lent (seulement un oeuf produit par an), choisir un bon partenaire est essentiel pour le succès reproducteur de l'individu. Ainsi on évitera de s'apparier avec un proche parent ou un individu au génome proche. Avoir une descendance porteuse d'une grande diversité génétique signifie en effet éviter les allèles récessifs provoquant malformations et maladies, et une meilleure résistance aux maladies.
Les oiseaux semblent capables d'évaluer la similarité du génome de leur partenaire avec le leur, et les femelles cherchent à s'accoupler à d'autres mâles que leur partenaire si cette similarité est trop grande. Cela a notamment été démontré chez le Bruant des prés, dont les femelles cherchent à s'accoupler avec les mâles porteur d'un complexe majeur d'histocompatibilité (Mhc) différent du leur. Le Mhc est un ensemble de gènes dont le rôle est essentiel pour le système immunitaire des vertébrés. Il est notamment avantageux pour un individu de posséder un Mhc très variable, car cela lui conférera une résistance à un large panel de pathogènes. Des suivis sur plusieurs années ont montré que les femelles des bruants associées à un mâle au Mhc semblable au leur vont certainement copuler avec d'autres mâles, tandis que celles en couple avec un mâle au Mhc suffisamment différent se contenteront de ce partenaire. Bien que l'étude n'ait pu mettre en évidence la façon dont les femelles discriminait le Mhc des mâles, il est raisonnable de penser que l'olfaction intervient dans ce processus, comme cela se fait chez les mammifères et poissons, en attendant de le voir confirmé ou infirmé par d'autres expériences plus ciblées.
En savoir plus
http://nationalzoo.si.edu/ConservationAndScience/MigratoryBirds/Science_Article/default.cfm?id=36
http://news.nationalgeographic.com/news/2003/05/0527_030527_birdscent.html
Blomqvist D., Andersson M., Küpper C., Cuthill I.C., Kis J., Lanctot R.B., Sandercock B.K., Székely T., Wallander J. and Kempenaers B., 2002. Genetic similarity between mates and extra-pair parentage in three species of shorebirds. Nature 419: 613-615
Freeman-Gallant C.R., Meguerdichian M., Wheelwright N.T. and Sollecito S.V., 2003. Social pairing and female mating fidelity predicted by restriction fragment length polymorphism similarity at the major histocompatibility complex in a songbird. Molecular Ecology 12: 3077-3083
Nevitt G.A., 2008. Sensory ecology on the high seas: the odor world of the procellariiform seabirds. The Journal of Experimental Biology 211: 1706-1713 (article)
Steiger S.S., Fidler A.E., Valcu M. and Kempenaers B., 2008. Avian olfactory receptor gene repertoires: evidence for a well-developed sense of smell in birds? Proceedings of the Royal Society B 275(1649): 2309-2317 (article)
{kind=link}